Introduction
The prevention of age-related degeneration and diseases such as, cancer, diabetes, liver disease, stroke and neurodegeneration, continue to dominate medical research [1,2]. Oxidative stress has been implicated as a key contributing factor in the development of these conditions [3,4]. It has been shown that high levels of reactive oxygen species (ROS) causes damage to cellular proteins, membrane lipids and nucleic acids, leading eventually to cell death and the onset of disease [5]. Thus, in order for our bodies to effectively respond to oxidative stress and restore a “normal” cellular redox balance, an up-regulation of these low-molecular-weight antioxidants and protective enzymes must occur [6].
Studies investigating the use of edible seaweeds to regulate biological systems have grown in number. Of these, Sea tangle (Laminaria japonica), an age-old Pacific and Asian food resource - typically consumed for its high contents of dietary fiber, carbohydrates, minerals and protein - has attracted growing interest for its bioactive potency and prophylactic activity [7,8]. Previous studies have supported the benefits of sea tangles for degenerative chronic diseases through antioxidant effects [9,10], as well as its hypotensive effects [11], anti-hyperlipidemic and anti-atherosclerotic activities [12].
In order to increase the GABA (gamma amino butyric acid) contents within sea tangle and elicit significant antioxidant effects, recent studies have implemented a specified fermentation process using Lactobacillus brevis BJ20. The results of these studies have indicated that superoxide radical scavenging, catalase (CAT) and glutathione peroxidase (GPx) activity were enhanced, whereas the lipid peroxidation level was diminished following fermented sea tangle (FST) supplementation [13, 14].
It is well documented that another, particularly damaging, effect of ROS and RNS is on DNA structure and function; typically resulting in base modification [15], single and double-strand breaks [16], damage to the DNA repair system [17], and associated with carcinogenesis [4]. Most damage is caused by the hydroxyl radical (OH·), which is considered the most powerful oxidizing agent in biological systems due to its very high reactivity and short half-life [18]. The presence of other reactive species, such as superoxide (O2-) and hydrogen peroxide (H2O2) also provide a source for other reactive intermediates that can collectively result in the production of hydroxyl radicals. To our knowledge, only one previous study has evaluated the antioxidant effects of FST within a clinical trial setting [13], and no studies have yet reported its possible protective provision for DNA. This coupled with the growing evidence that antioxidant capacity decreases with age [19], forms the premise of the current study, which aims to investigate the effects of FST supplementation on antioxidant capacity in aged women.
Methods
Subjects
Forty, moderately active, senior women (72.8 ± 5.0yrs) participated in a randomized, double-blind and placebocontrolled study (Table 1). Two groups were formed, one placebo group (73.6 ± 4.6 yrs) and one FST group (72.0 ± 5.5yrs). Subjects suffering with hypertension, diabetes, chronic degenerative disease (arthritis), cardiovascular disease, and/or obesity were excluded from the study. Participants were also excluded if they experienced adverse reaction to supplementation prior to the experimental period or were unable to participate due to particular medication. All participants completed a written informed consent to partake in this study, which was approved by Kyungsung University Ethics Committee (KSU-16-05-001).
Material preparation and treatment
FST was prepared following a recently modified procedure [20]. Sea tangle was added to water at a ratio of 1:15 (w/v) with the addition of yeast extract and glucose, based on the amount of sea tangle added to the mixture. After autoclaving at 121 °C for 30 min, a sea tangle solution was obtained and the culture broth (Lactobacillus brevis BJ20) was added to the solution at a concentration of 1.2% (v/v) (Table 2), which was mixed and incubated at 37 °C for 2 days (Figure 1).
Based on a previous clinical study [20], the content of GABA within FST was controlled to range between 40~60 mg within 1,000 mg of FST using high performance liquid chromatography analysis during the preparation process. Participants ingested either 1,000 mg of FST (FST) per day or 1,000 mg sucrose placebo (CON) per day, for 6 weeks. Subjects were asked to abstain from any regular exercise. Fasting venous blood samples were taken before (pre-experiment) and after (post-experiment) the 6 weeks’ period. Collected blood samples were clotted for 30 min and centrifuged for 15 min at 1,000 ×g to separate into serum, and stored at -80°C and analyzed by immunoassay.
Measurements
Serum SOD (Superoxide dismutase) level was determined using a commercially available kit, Cayman’s Superoxide dismutase assay kit (Cayman Chemical Company, Ann Arbor, MI, USA). Tetrazolium salt was utilized to detect superoxide radicals generated by xanthine oxidase and hypoxanthine, which react to form a yellow formazan dye. The detection limit of the assay was 0.025-0.25 U/ml SOD. Inter and intra-assay coefficients of variation were 3.7 and 3.2%, respectively. Absorbance was measured at a wavelength of 440-460 nm and enzyme activity (U/ml) was calculated.
Serum GPx (Glutathione peroxidase) level was measured using the Cayman GPx Assay Kit (Cayman Chemical Company, Ann Arbor, MI, USA). The Co-substrate mixture contained NADPH, glutathione, and glutathione reductase, and the reaction was initiated by adding 20 μl of cumene hydroperoxide. One unit of GPx was defined as the amount of enzyme that catalyzes the oxidation of 1.0 nmol of NADPH per minute at 25°C. Absorbance was measured at a wavelength of 340 nm and enzyme activity (U/ml) was calculated.
Serum GSR (Glutathione reductase) level was measured using the commercially available Cayman Chemical Glutathione Reductase Assay Kit (Cayman Chemical Company, Ann Arbor, MI, USA), which measures the rate of NADPH oxidation. The reaction was initiated by adding 50 μl of NADPH and this is accompanied by a rapid decrease in the absorbance at 340 nm. One unit of GSR activity was calculated as the amount of enzyme that catalyzes the oxidation of 1.0 nmol of NADPH per minute at 25°C
Serum TBARS (thiobarbituric reactive species) content was determined with the OxiSelectTM TBARS Assay Kit, a commercial kit provided by Cell Biolabs Inc. (San Diego, CA, USA). The malondialdehyde containing samples were first reacted with TBA at 95°C. After a brief incubation, the malondialdehyde protein adducts content in the serum was determined by comparison with a predetermined malondialdehyde standard curve. Spectophotometric measurement was used to calculate serum TBARS content (μM) at 532 nm absorbance.
Oxidative stress marker to DNA, 8-OHdG (8-hydroxodeoxyguanosine) was measured using a competitive enzyme-linked immunosorbent assay kit from JaICA (Japan Institute for the Control of Aging, Shizuoka, Japan), following manufacturers’ instructions. This test used a monoclonal antibody N45.1. Absorbance from the wells was measured at 450 nm with a microtiter plate reader.
Statistical Analysis
All statistical analyses were conducted with SPSS Windows version. 21.0. In order to verify the impact of FST supplementation, two-way analysis of variance (ANOVA) with repeated measure was conducted to assess the main effects between groups. Descriptive statistics were used to report on each variable. Statistical significance was set at α = 0.05.
Results
Prior to the experimental period the baseline values of serum SOD in the FST and placebo group were 5.56 ± 0.35 U/ml and 5.70 ± 0.47 U/ml, respectively. As shown in Figure 2A, SOD levels in the FST group (6.15 ± 0.46 U/ml) tended to increase, whereas the placebo group (5.29 ± 0.55 U/ml) tended to decrease. But there was no significant difference in main effect between groups (FST treatment) (p=.353) or within groups (time, 6-week period) (p=743) in serum SOD levels. However, there was a significant interaction effect between FST treatment and time in serum SOD (p<.01).
Following treatment, GPx levels in the FST group increased (195.57 ± 43.96 nmol/min/ml to 217.73 ± 56.95 nmol/min/ml), whereas the placebo group tended to decrease (190.89 ± 43.02 nmol/min/ml to 182.29 ± 38.94 nmol/min/ml) (Figure 2B). Statistically, GPx level in the FST group was not significantly different compared to that of the placebo group following the 6-week supplementation period (p=.055). However, there is no significance within groups (time, 6-week period) (p=.051)
GSR levels in serum tended toward an increase in the FST group (179.73 ± 22.73 nmol/min/ml to 195.751 ± 18.63 nmol/min/ml), while conversely, a slight decrease was recorded in the placebo group (171.79 ± 25.19 nmol/min/ml to 158.14 ± 35.55 nmol/min/ml) (Figure 2C). Statistically, the main effect of FST supplementation on serum GSR levels was significant such that the FST group had higher levels than the placebo group following the 6-week treatment (p<.01). However, there is no significance within groups (time, 6-week period) (p=.450).
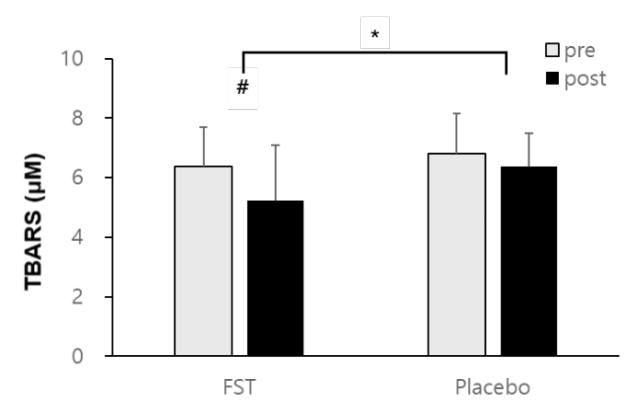
The results of this study showed a reduction in TBARS values in both the FST group (6.58 ± 0.84 μM to 5.67 ± 0.65 μM) and placebo group (6.89 ± 0.72 μM to 6.71 ± 1.35 μM) (Figure 3). The between groups difference following 6 weeks of FST supplementation (p<.05) and within groups before and after the treatment (p<.05) were significant. Whereas, there was no significant interaction effect between FST treatment and time in serum TBARS level (p=.273).
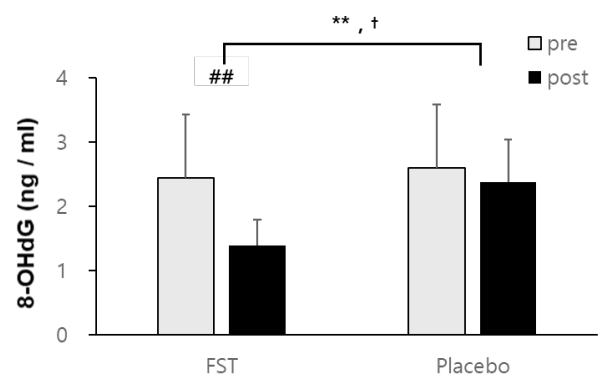
In regard to oxidative damage to DNA, 8-OHdG levels were significantly decreased within FST (2.39 ± 0 .44 ng/ml to 6.89 ± 0.72 ng/ml) whereas placebo groups were not statistically changed (2.57 ± 0.40 ng/ml to 2.46±0.56 ng/ml) (Figure 4). These resulted in a significant difference in both between groups (FST supplementation) (p<.01) and within group (time) (p<.01) after the 6-week treatment. A significant interactive effect of FST treatment and time (p<.05) was also demonstrated in serum 8-OHdG levels.
Discussion
Sea tangle, L. japonica, has been investigated for its numerous health preserving and enhancing effects [9-12], including its antioxidant properties [13-14]. The current study demonstrated that FST supplementation for a 6-week period enhanced the serum level of SOD and GSR (Figure 2), while reducing biomarkers of lipid peroxidation (TBARS) and oxidative DNA damage (8-OHdG) (Figure 3, 4). These results have shed greater light on the antioxidant potential of FST, supporting initial investigations suggesting a protective effect against lipid peroxidation [12] and an enhancement in the enzymatic antioxidant defense system [7, 13], thereby combating the cytotoxic oxidants that mediate the pathogenesis of many chronic diseases.
The cells’ main defense mechanism against oxygen free radicals includes enzymes such as SOD, GSR and GPx, among other low molecular weight antioxidants and antioxidant nutrients. Lee et al. [13] reported that FST quenched O2-radicals; while Kang et al. [14] went on to demonstrate a significant augmentation in SOD activity following 4 weeks of FST supplementation. It is of particular importance that the present study corroborated these results in senior subjects, whose aging cells grow increasingly inefficient at eliminating oxidative stressors seen particularly in the mitochondrion, where there is increased ROS production and even damaged membrane integrity with age [21]. The scavenging of O2- by SOD will reduce OH· production and potential site-specific biological damage to lipid membranes, proteins and DNA [22].
Though there is a reactive byproduct of SOD in the form of H2O2, it may be effectively detoxified to water by GPx in a reduction reaction involving glutathione (GSH) [23]. Reduced GPx activity has been identified as a predictor of increased cardiovascular risk [24, 25]. Six weeks of FST administration in this study, however, did not lead to the significant enhancement of GPx level (Figure 2B). These results are supported by a previous study [15] in which FST ingestion for 4 weeks showed no significant effect on GPx activity. However, a longer experimental period may have rendered differing by FST supplementation [20]. Taken together, a longer treatment period, potentially 8 weeks, may be required to demonstrate an enhancement of serum GPx levels in the older women.
Tight regulation of GSH, and its oxidized form (GSSG), is imperative to cell survival; so much so that an imbalance in GSH has been linked to a wide range of pathologies, including Alzheimer’s disease, aging [26], and cancer [27]. GSR works hand-in-hand with GPx to help regulate the redox state of GSH (GSH : GSSG ratio), including decomposition of H2O2. Thus, reducing levels of GSR reported in the latter stages of life [28], particularly concerning to the aging population. Our study demonstrated a significant difference in GSR activity between the FST and CON groups (Figure 2C). GSH is a vital scavenger, recycled by GSR to combat oxidative stress, therefore an up-regulation of GSR via dietary intervention would aid in optimizing its redox state, and thus reduce deleterious oxidative damage.
Cellular membranes are especially susceptible to oxidation due to their high concentrations of polyunsaturated fatty acids [29]. In consistent with previous findings [7, 15, 16], we showed that fermented L. japonica intake had a significant ameliorative effect on lipid peroxidation, lowering TBARS (Figure 3). TBARS, thiobarbituric reactive species is the end product of whole-organism lipid peroxidation [30]. If the deleterious modification of cellular membranes can be alleviated by dietary supplementation, FST may provide a viable defense against lipid-peroxidation-associated conditions such as arteriosclerosis, neurodegenerative disease, and cardiovascular disease [31].
Research has not been undertaken concerning the antioxidant effects of FST supplementation on oxidative damage to DNA, to our knowledge. Thus, the present study recorded the changes in the DNA-oxidized adduct, 8-OHdg following FST supplementation. A significant protective effect against free radical-induced DNA damage was observed in the FST group, compared to the CON (Figure 4). This finding is important, as 8-OHdg is one of the predominant forms of free radical-induced oxidative lesion and widely considered a critical biomarker of oxidative stress and carcinogenesis [32]. Increased 8-OHdg has also been implicated in both nuclear and mitochondrial DNA damage, associated with age [33]. Therefore, FST may be considered an effective and versatile dietary source of antioxidants, capable of providing DNA protection, particularly antimutagenic effects.
Conclusions
The present study highlighted the widespread antioxidant effects of FST in an elderly population. The upregulation of antioxidant enzyme activity (SOD and GSR) and inhibition of lipid- and DNA-oxidative damage, suggests that FST may be an effective nutritional countermeasure against the age-related increase in oxidative stress.
Considering that some antioxidants, in excessive levels, may be potentially hazardous (e.g., toxicity in humans and DNA oxidation in leukocytes) [34], further research is needed concerning the impact of FST and other dietary interventions on the antioxidant defense systems in specific biological sites in the aging population.